
紫檀烷和异黄烷是豆科植物中广泛存在的重要次生代谢产物,具有抗肿瘤、美白、抗菌、抗氧化等多种药理活性,也是植保素。紫檀烷还原酶(pterocarpan reductase, PTR)是催化紫檀烷发生C-O键断裂转化为异黄烷的关键酶,其催化机制尚不明确。高效PTR酶的发现及催化机制研究,对于异黄烷类活性化合物的绿色合成具有重要意义。近期,北京大学-云南白药国际医学研究中心副主任叶敏教授团队从乌拉尔甘草中鉴定六条紫檀烷还原酶(GuPTR1-6),并解析了GuPTR1/medicarpin/NADP+复合物晶体结构(1.8 Å)。基于结构分析与定点突变,揭示了紫檀烷还原酶的催化机制。

1. 甘草中紫檀烷还原酶的鉴定
通过本地Blast筛选获得候选基因后,研究者从甘草(Glycyrrhiza uralensis Fisch.)中克隆得到六个PTR基因 (GuPTR1-6)。通过体外酶活实验验证,研究者发现六条PTR均能催化美迪紫檀素(1)开环生成驴食草酚 (1a),如图1所示。
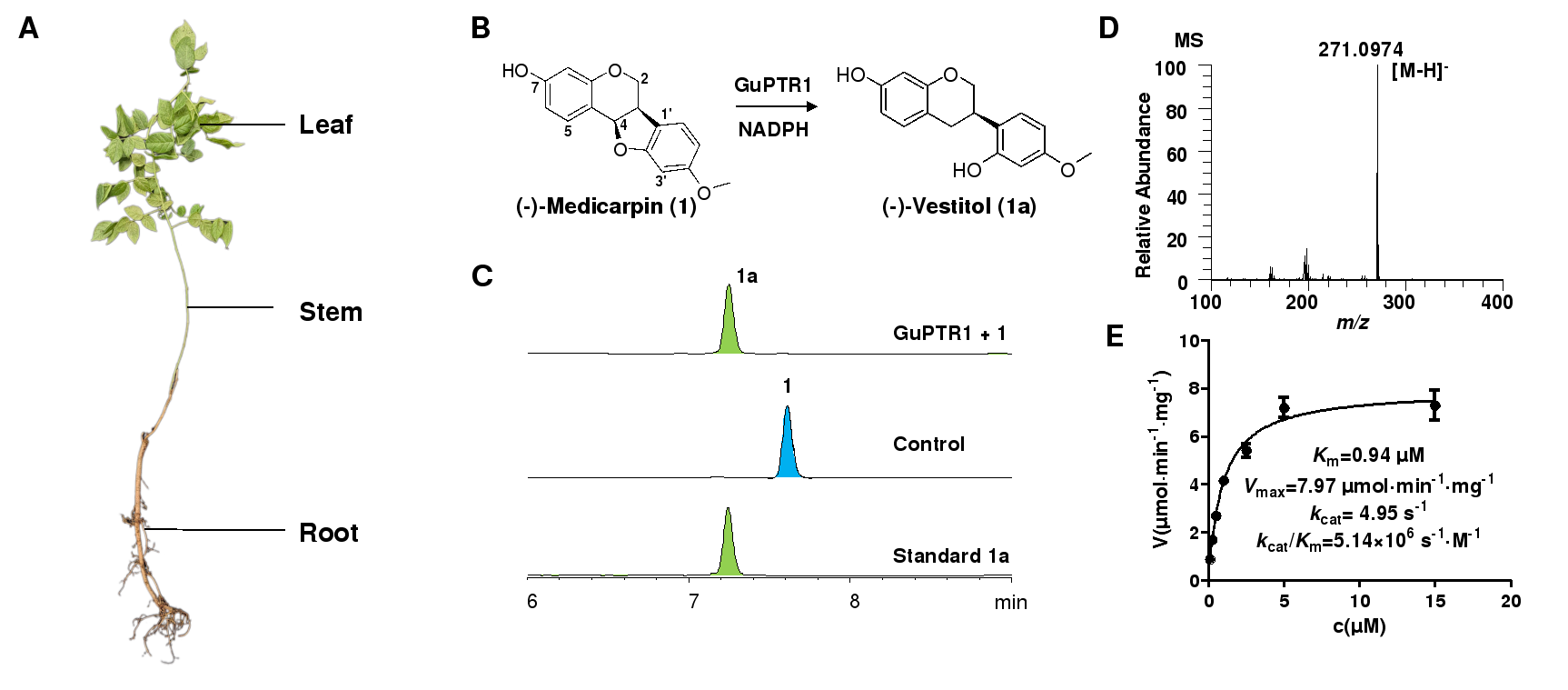
图 1. GuPTR1的功能表征
2. 紫檀烷还原酶特异性识别具有7-OH的底物
通过底物谱考察,研究者发现这些PTR酶仅能催化具有7-OH的紫檀烷,而对7-OH被取代的底物均不能催化(图2)。该结果提示,尽管7-OH距离底物的反应中心较远,其在PTR的催化过程中发挥重要作用。
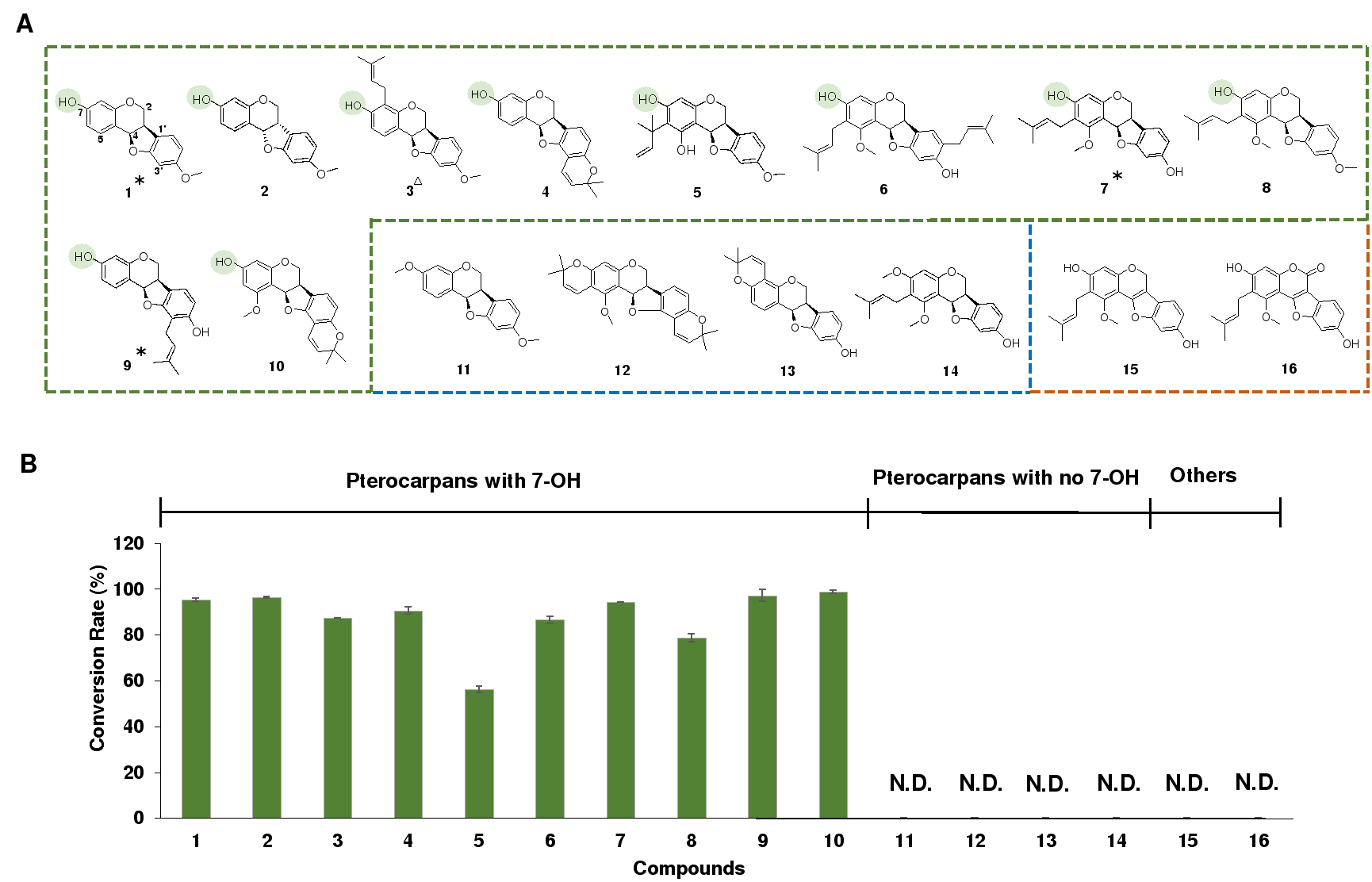
图2. GuPTR1的底物谱考察
3. PTR的晶体结构与催化机制研究
为了探究紫檀烷底物7-OH在PTR催化过程中发挥的作用,研究者培养并解析了GuPTR1与美迪紫檀素及NADP⁺的复合物晶体,是首个PTR酶的晶体结构。通过定点突变、分子动力学模拟等手段,发现K135对GuPTR1的催化活性至关重要,其可能介导7-OH脱质子,促进芳环体系发生电子重排,导致C-O键断裂形成醌式中间体。随后,美迪紫檀素C-4位接受来自NADPH的氢,促使芳香环体系重建(图3)。
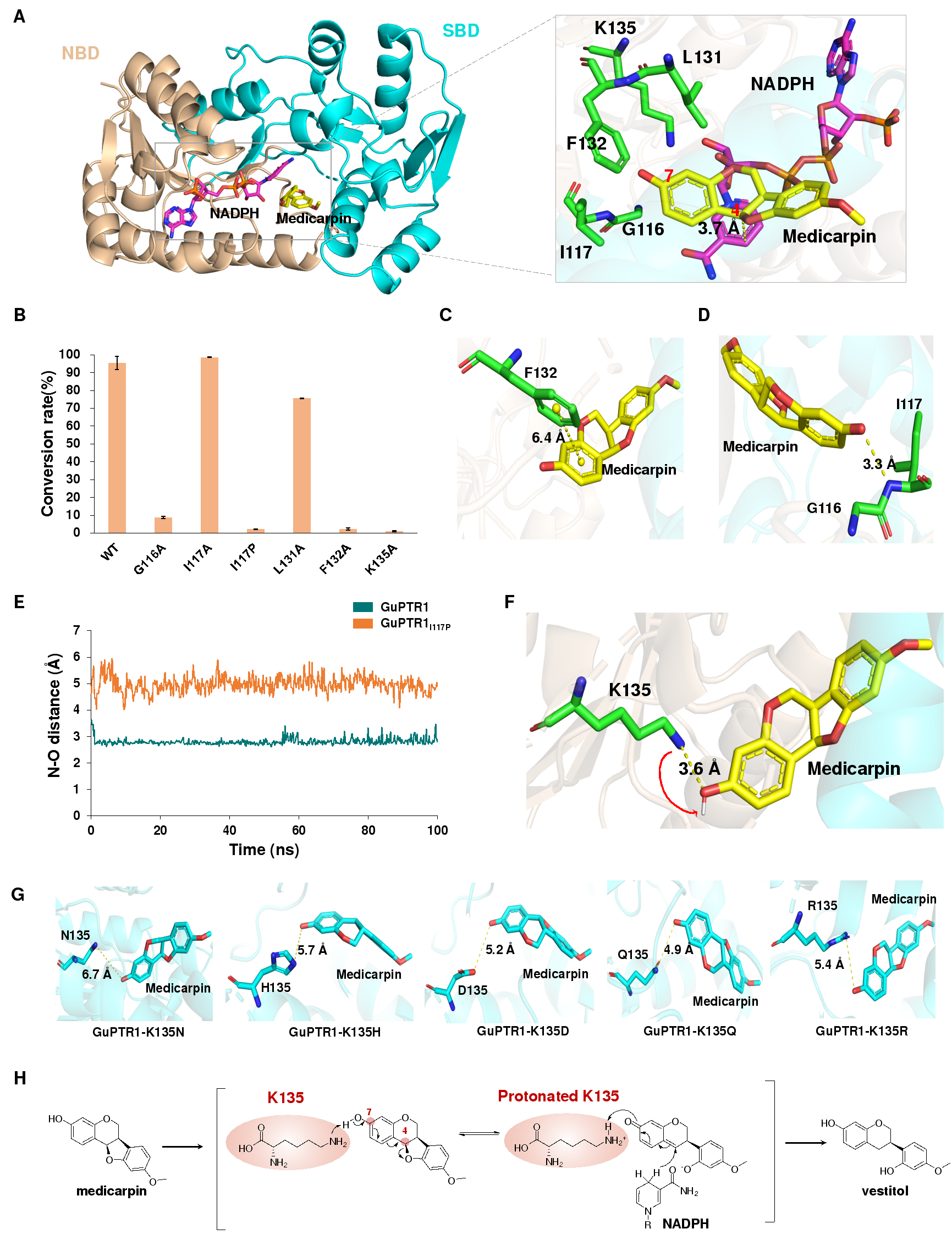
图 3. GuPTR1 的催化机制
4. K135在植物界PTR中十分保守
为了进一步探究K135在PTR催化中的重要性,本研究将甘草来源的GuPTR2-6中的K135突变为丙氨酸,发现催化活性显著降低(图4A)。进一步从CNGB数据库中获取了100条植物PTR序列,发现该K135在其中99条PTR中存在(图4B)。随机选择具有K135的TpPTR1、MnPTR、TcPTR进行功能验证,发现三者均能高效催化底物转化,而缺少K135的TpPTR2则活性微弱。定点突变实验进一步说明该保守赖氨酸残基是PTR酶家族催化功能不可或缺的关键位点(图4 C)。
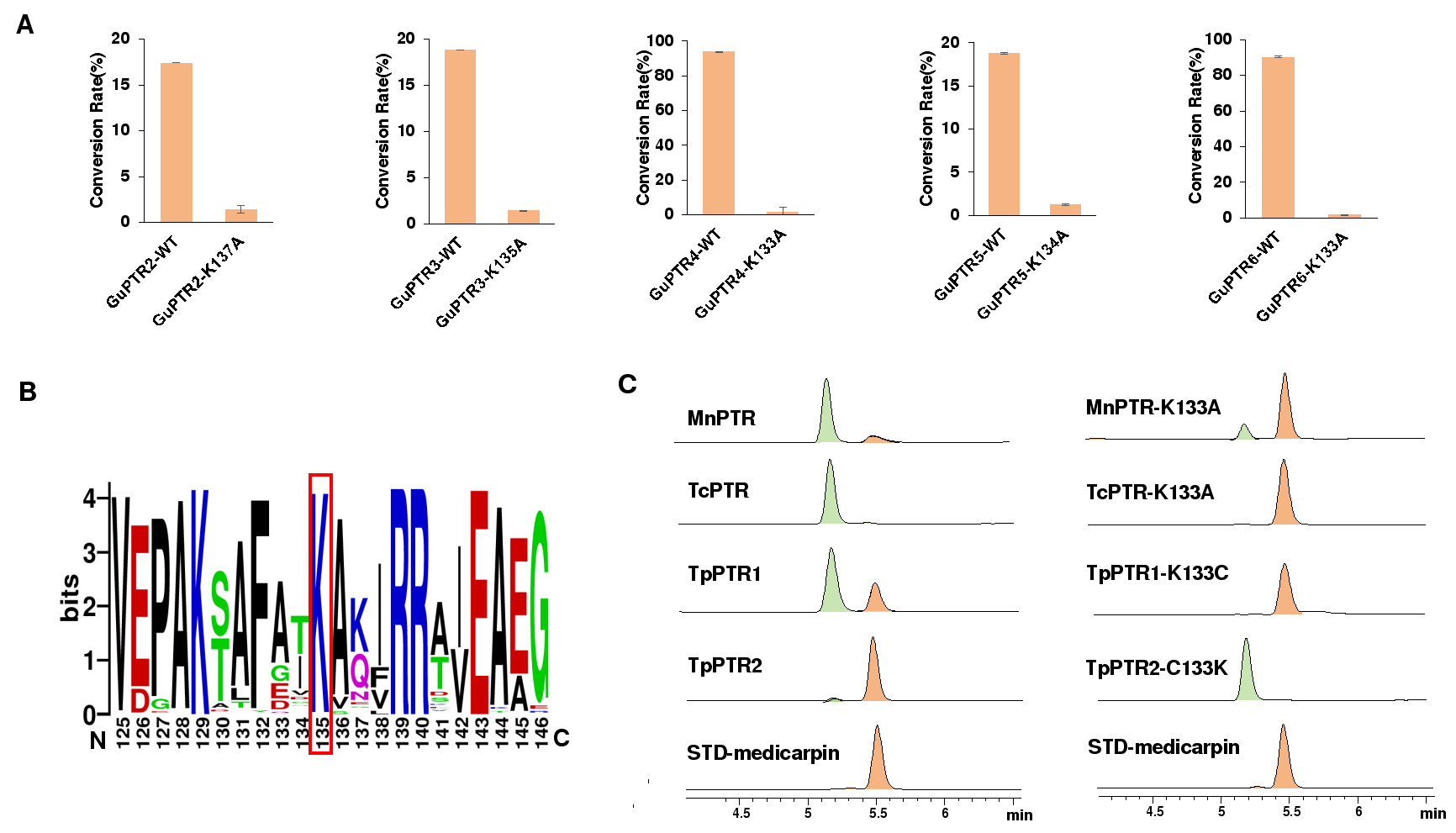
图 4. 赖氨酸残基在 PTR 酶催化反应中的作用
5. 祖先酶N0的重建与应用
根据系统发育分析,PTR与 PLR (松脂醇-落叶松脂醇还原酶)和PCBER(苯基香豆烷苄基醚还原酶)在进化上高度关联,且均能催化具有4-(呋喃-2-基)苯酚结构单元的底物,提示三者很可能具有类似的催化机制。研究者从CNGB数据库获得了100条PLR与100条PCBER,与之前获得的100条PTR共同构建了系统发育树,并重建了三者的共同祖先酶N0(图5A)。N0能够催化三种不同类型的4-(呋喃-2-基)苯酚衍生物(紫檀烷、松脂醇和苯基香豆烷苄基醚)发生C-O键断裂,是一种普适性催化剂(图5B-D)。
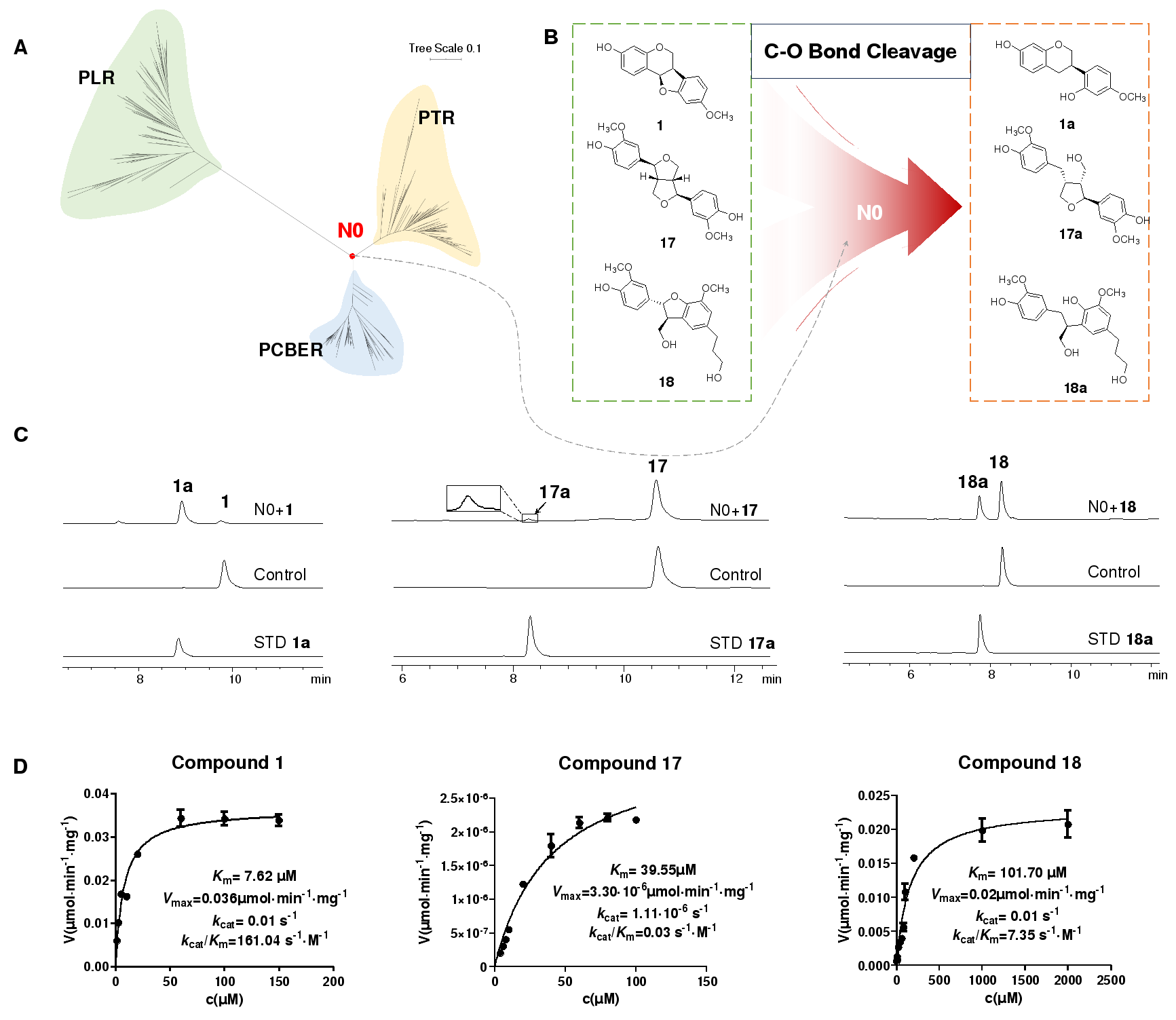
图 5. 祖先酶N0的重建与催化功能
综上,该研究从药用植物甘草中鉴定了六条紫檀烷还原酶(GuPTR1-6),并解析了GuPTR1/medicarpin/NADP+复合物晶体结构(1.8 Å)。基于结构分析与定点突变,该研究揭示了紫檀烷还原酶的催化机制。其中,高度保守的赖氨酸K135介导7-OH脱质子,进一步通过共轭体系中的电子传递触发C-O键断裂。该催化机制同样适用于PLR (松脂醇-落叶松脂醇还原酶)和PCBER (苯基香豆烷苄基醚还原酶)等其他催化4-(呋喃-2-基)苯酚衍生物C-O键断裂的生物合成酶。该研究还通过祖先序列重构,获得了能够催化三类4-(呋喃-2-基)苯酚衍生物开环的多功能还原酶N0,为该类反应的绿色催化提供了新的酶学工具。
北京大学药学院天然药物及仿生药物全国重点实验室、北京大学-云南白药国际医学研究中心叶敏教授为论文的通讯作者。2021级直博生李鸿晔、2020级长学制学生邹建霖为论文的共同第一作者,博士后张梦、赵春雪等做出了重要贡献。该研究工作得到国家重点研发计划、国家自然科学基金项目的支持。

叶敏教授团队合影